|
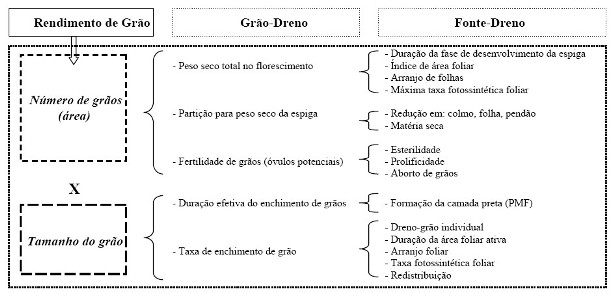
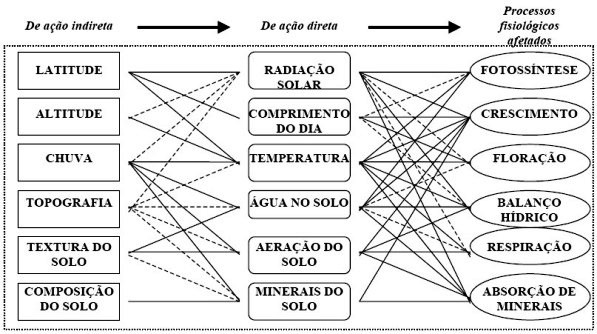
2. A planta e a plantaçăo de milho O milho (Zea mays L.) apresenta características fisiológicas favoráveis ŕ eficięncia de conversăo do carbono mineral em compostos orgânicos (Williams et al. 1965; Gifford & Evans, 1981). A planta de milho é considerada um sistema eficiente para converter a energia luminosa em matéria seca do grăo, se bem que esse enfoque de incremento do rendimento do grăo, pode estar negligenciando outras partes da planta economicamente importantes. O genótipo influencia a quantidade de energia fixada, bem como sua distribuiçăo para as várias partes da planta. O rendimento de grăo em uma dada área de terreno é considerado em seus componentes número de grăos por unidade de área x tamanho (e peso) de grăo. A melhoria da eficięncia do rendimento de grăos de milho pode ser observada no esquemático apresentado na Figura 1.
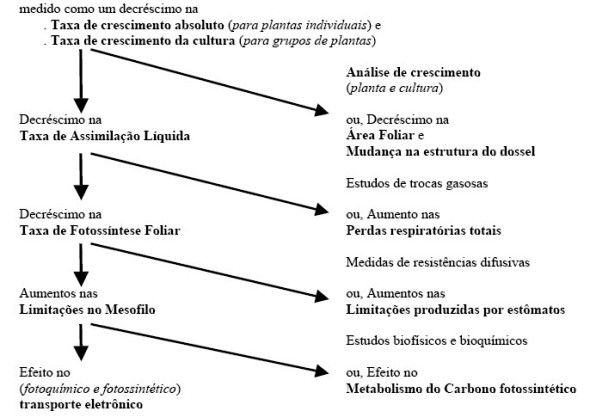
Alguns processos fisiológicos săo envolvidos para determinar esses componentes, e devem ser explorados para mudanças genéticas, visando melhoria do rendimento (Figura 2).
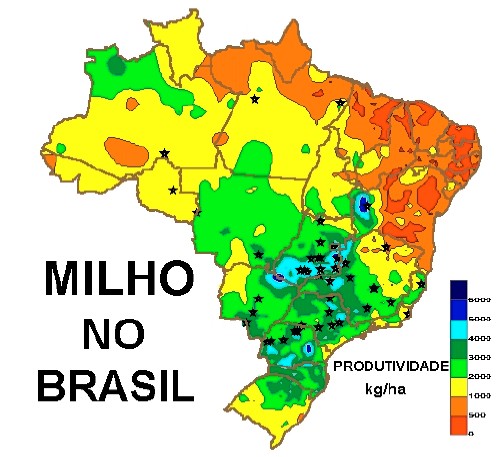
O número de óvulos presentes na época do florescimento representa o número de grăos potenciais que, entretanto, podem ser reduzidos por eventos posteriores ao florescimento (Derieux et al., 1983). Segundo Fuchs (1968), citado por Derieux et al. (1983), existe uma relaçăo entre o número de óvulos e precocidade. Os resultados indicam que número de óvulos inicial é altamente correlacionado com a superfície foliar no início da diferenciaçăo e que genótipo influenciou mais que o fator ambiental temperatura. Segundo Evans (1975) o período de fertilizaçăo até rápido acúmulo de matéria seca no grăo (lag phase), pode variar de 12 para 20 dias em cultivares de milho tropical, em contraste com cereais de grăos pequenos, tais como trigo, que tem uma lag phase de menos de sete dias. Grăos que abortam săo normalmente os mais jovens, no topo do sabugo da espiga, devido ao rápido acúmulo de matéria seca pelos grăos mais velhos abaixo no sabugo. Aborto ocorre a despeito da relativamente alta concentraçăo de açúcares solúveis em colmos e grăos. Milho tem potencial morfológico para produzir mais de uma espiga, e os mecanismos que controlam prolificidade săo provavelmente relacionados com aqueles envolvidos em esterilidade. Evidęncias experimentais apontam que aborto de espiga está relacionado com competiçăo intraplanta por fotoassimilados, durante o período crítico de crescimento. Diferenças de rendimento de milho podem refletir o nível de práticas agronômicas nas diferentes regiőes. Problemas de fertilidade do solo, doenças de plantas, insetos, plantas daninhas, disponibilidade de umidade, e uma tendęncia para os extremos no clima em áreas tropicais; além de cultivos de subsistęncia, em condiçőes precárias ou em associaçăo com outros cultivos, podem explicar a variaçăo nos rendimentos, em geral baixos. Aumentos no rendimento podem ocorrer devido ŕ melhor administraçăo agroeconômica dos cultivos; entretanto, em patamares de produtividade mais elevados, os incrementos no rendimento devem passar pelo melhoramento genético, objetivando produtividade e/ou tolerância aos estresses. Em áreas tropicais, a maioria dos altos rendimentos săo restritos em áreas intermediárias ou de alta altitude, tendo longas estaçőes chuvosas. Segundo Fisher & Palmer (1983), citando vários autores, rendimentos de 12 t.ha-1 tem sido obtidos em latitude 18ş S, altitude 1.500m. Em terras baixas tropicais, rendimentos podem atingir 5 a 8 t.ha-1, com boa administraçăo. Em regiőes temperadas, rendimentos máximos próximo de 20 t.ha-1 săo relatados. Rendimentos comerciais de 10 t.ha-1 săo comuns (Frey, 1971). Goldsworthy et al. (1974) apontam que produçăo de fitomassa total em ambientes tropicais năo é um fator limitante para o rendimento de milho. Yamaguchi (1974) e Goldsworthy et al. (1974) atribuíram o baixo rendimento de grăos, em regiőes tropicais, ŕ deficiente partiçăo da matéria seca para o grăo. Em áreas tropicais abaixo de 1.000 m de altitude, a planta de milho acumula considerável matéria seca no colmo, bainhas de folhas, sabugo e brácteas após florescimento. Parte dessa matéria seca é translocada para o grăo mais tarde, no período de enchimento de grăos (Palmer et al., 1973). Resultados experimentais relatados por Goldsworthy et al. (1974) apontam que o rendimento de milho é limitado principalmente pelo tamanho do grăo-dreno, embora sob algumas perdas ambientais de área foliar (fonte) próximo da maturidade pode reduzir o rendimento de grăos. Milhos tropicais geralmente tem maturaçăo tardia, porte alto e grande pendăo. Eles mostram um nível maior de dominância apical, baixo índice de colheita, e, săo menos eficientes em retranslocar para os grăos a matéria seca previamente depositada no colmo. Tais características tęm provável importância para a adaptaçăo do milho em ambientes tropicais, em competiçăo vantajosa com plantas de rápido crescimento, e minimizado efeito da perda de folhas por insetos. Ou seja, milho tropical tem abundância de folha (fonte) e possivelmente tem mais problemas de partiçăo e dreno (grăo) do que milhos temperados. Reservas de açúcares no colmo pode ser, evolutivamente, resultado da tolerância a seca e pode ser associada com resistęncia para podridőes do colmo. Em altitudes superiores a 1.000m, os cultivares de milho tropical apresentam a duraçăo do crescimento mais longa que em terras baixas tropicais e é limitado ou por baixas temperaturas ou disponibilidade de umidade para plantio (Wilson et al., 1973). Embora os rendimentos em terras altas tropicais sejam maiores que aqueles em terras baixas tropicais, Goldsworthy & Colegrove (1974) e Yamaguchi (1974) concluíram que rendimentos em ambas as áreas foram limitadas pela capacidade do grăo-dreno em acumular fotoassimilados. Algumas dessas diferenças em partiçăo săo devidas ao ambiente e interaçăo genótipo x ambiente. Entretanto, há consideráveis variaçőes genéticas quando diferentes genótipos crescem em um mesmo ambiente. Por causa da sensibilidade ao fotoperíodo de genótipos tropicais, especialmente para faixas de latitudes superiores a 33ş Sul, uma comparaçăo năo pode ser feita em ambientes temperados. Segundo Badu-Apraku et al. (1983), uma reduçăo no rendimento está associada com o período de enchimento de grăos mais curto. Durante um rápido enchimento de grăos, o aumento na matéria seca do grăo resulta da utilizaçăo dos efeitos combinados de assimilados temporariamente estocados em partes vegetativas da planta e produzidos através da fotossíntese. Em altos regimes de temperatura diurna/noturna a acumulaçăo de matéria seca durante o enchimento de grăos é reduzida. Rendimentos de grăos por planta também é menor sob altas temperaturas. Diferenças no rendimento de grăos năo săo sempre causadas por diferenças na duraçăo do cultivo, medidas em dias ou em acumulaçăo de unidades de calor. Nas regiőes tropicais, onde é menor a variaçăo de temperatura, umidade é fator preponderante. A produçăo é influenciada pela disponibilidade de água no solo, especialmente na fase que antecede ao pendoamento (Mundstock, 1970). 2.1. Produçăo de milho para altas performances e rendimento de grăos O rendimento de grăos de milho é determinado basicamente pelo número de grăos por unidade de área e, em menor escala, pela massa individual do grăos (Richards, 2000). Vide também Figura 1, que mostra esquemático de melhoria da eficięncia do rendimento de milho. A melhoria do rendimento em milho tem sido associada com o aumento na tolerância a estresses (Duvick, 1992, 1997; Tollenaar & Wu, 1999, Tollenaar et al., 2001) e o aumento na tolerância a estresses podem resultar, em parte, da seleçăo para aumento da estabilidade do rendimento (Durăes et al., 2002). 2.2. Produçăo de milho sob condiçőes ambientais sub-ótimas (estresses abióticos e bióticos) Os rendimentos de grăos e outras variáveis de interesse para a caracterizaçăo da performance adaptativa dos genótipos de milho de diferentes grupos de maturidade (normal, precoce e superprecoce), cultivados em diferentes locais das regiőes Centro e Sul do Brasil, săo apresentados na Tabela 1, por Durăes et al. (2002); e, os dados permitem observar variaçőes significativas no rendimento de grăos, quando os ensaios foram separados por grupos de altitude (≥700 e <700m), visualizados também nas Figuras 3 e 4. No geral, os maiores rendimentos foram obtidos na regiăo Sul, e em genótipos de grupos de maturidade tendendo de precoce a superprecoce. Na regiăo Centro, as maiores produtividades foram obtidas pelos genótipos de grupo de maturidade precoce. Em ambas as regiőes observaram-se os maiores rendimentos em ensaios conduzidos em locais de altitudes ≥700m, sendo que na regiăo Sul foram apresentadas as maiores instabilidades de rendimento em relaçăo ŕ regiăo Centro, para genótipos de milho dos tręs grupos de maturidade.
Para ambas as regiőes, Centro e Sul, e grupos de maturidade (Normal, Precoce e Superprecoce) a produtividade (kg/ha de grăos) seguiu a tendęncia decrescente para os materiais com background genético (Híbrido Simples, Híbrido Triplo e Variedade), respectivamente. Segundo Durăes et al. (2002), este mesmo comportamento, embora mais disperso, foi observado para o Índice de Estresse Ambiental (bi), com base na produtividade de genótipos em diferentes altitudes (≥700 e <700m). Tanto para produtividade quanto para Índice de Estresse Ambiental, algumas variedades atingiram valores superiores ŕ media, o que demonstra boa estabilidade de produçăo, inclusive em mais altos patamares de produçăo.
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||